Abstract : Since its discovery in 1923, the parathyroid hormone (PTH), was thought to be the sole hormone capable of stimulating bone resorption, renal tubular calcium reabsorption, calcitriol synthesis, and urinary excretion of phosphate. However, in 1987, the PTHrP (PTH-related peptide), was demonstrated to share most of the biological actions of PTH through the activation of the same receptor. This receptor was cloned in 1992 and named PTH/PTHrP receptor or PTH-R1. Both, PTH and PTHrP bind with great affinity to PTH-R1 and stimulate a signal transduction system involving different G-proteins, phospholipase C, and adenylate cyclase. A third member of the PTH family, the TIP-39 (tuberoinfundibular peptide), binds and activates another PTH receptor (PTH-R2).
There is evidence for other PTH receptors, a PTH-R3, probably specific for PTHrP in keratinocytes, kidney, and placenta, and a PTH-R4 specific for C-terminal PTH fragments. Activating mutations in the PTH-R1 gene cause Jansen type metaphyseal chondrodysplasia, whereas inactivating mutations are responsible for Blomstrand type rare chondrodysplasia and enchondromatosis.
The renal and bone PTH-R1 expression is upregulated in vitamin D deficient rats and by endotoxin, interleukin-2, dexamethasone, T3, and TGF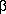 . On the contrary, PTH, PTHrP, angiotensin-II, IGF-1, PGE2, vitamin D, and chronic renal failure decrease its expression. In conclusions, the biological implications of the identification and cloning of different PTH receptors are at their beginning. The almost ubiquitous distribution of PTHrP and PTH-R1, the numerous PTHrP and PTH fragments, let us suppose the existence of other PTH-related receptors, and a great complexity of its potential actions on bone and mineral metabolism.
. On the contrary, PTH, PTHrP, angiotensin-II, IGF-1, PGE2, vitamin D, and chronic renal failure decrease its expression. In conclusions, the biological implications of the identification and cloning of different PTH receptors are at their beginning. The almost ubiquitous distribution of PTHrP and PTH-R1, the numerous PTHrP and PTH fragments, let us suppose the existence of other PTH-related receptors, and a great complexity of its potential actions on bone and mineral metabolism.
Key words: Parathyroid hormone, Parathyroid hormone-related peptide, Parathyroid Hormone receptor, Renal osteodystrophy, Review.
Resumen
Desde su descubrimiento en el aÒo 1923, la hormona paratiroidea (PTH) era la ˙nica capaz de estimular la resorciÛn Ûsea, la reabsorciÛn tubular de calcio, la eliminaciÛn renal de fÛsforo y la sÌntesis de calcitriol. Sin embargo, en el 1987 se identificÛ el PTHrP (PTH-related peptide), el cual ejerce la mayorÌa de las funciones biolÛgicas de la PTH a travÈs del mismo receptor. Ese receptor, PTH/PTHrP o PTH-R1, fue clonado en el 1992. Las dos molÈculas, PTH y PTHrP se ligan al Èl con gran afinidad y estimulan un sistema de transducciÛn que implica la activaciÛn de diferentes proteÌnas G, la fosfolipase C y la adenilciclasa. Un tercer miembro de la familia PTH, el TIP-39 (tuberoinfundibular peptide), se liga y activa otro receptor de la PTH, el PTH-R2.
TambiÈn, hay evidencias de la existencia de otros receptores de la PTH (PTH-R3, PTH-R4). Las mutaciones activadoras del PTH-R1 provocan la condrodisplasia met·fisaria de tipo Jansen y las mutaciones inactivadoras son responsables de otra condrodisplasia rara, la enfermedad de Blomstrand y de ciertas encondromatosis.
La expresiÛn del PTH-R1 aumenta en el riÒÛn y el hueso de ratas deficientes en vitamina D y en respuesta a las endotoxinas, la interleuquina-2, la dexametasona, la T3 y el . Al contrario, la PTH, el PTHrP, la angiotensina-II, el IGF-1, la PGE2, la vitamina D y la insufficiencia renal crÛnica la disminuyen. En conclusiÛn, las implicaciones biolÛgicas del clonaje de los receptores de la PTH son innumerables. El hecho que el PTHrP y el PTH-R1 sean ubicuos y el hecho que hay varios fragmentos circulantes del PTHrP y de la PTH, supone la existencia de otros receptores especÌficos de esos fragmentos y por supuesto que la acciÛn de esas molÈculas sobre el metabolismo mineral y Ûseo es mucho m·s compleja de lo imaginado.
Palabras claves : Hormona paratiroidea, pÈptido relacionado a la PTH o PTHrP, Receptor de la hormona paratiroidea, Osteodistrofia renal, RevisiÛn.
IntroducciÛn
La hormona paratiroidea (PTH) se fija y activa el receptor de type 1 (PTH-R1), el cual fue identificado y clonado por el equipo de H. Juppner en Boston en el aÒo 1990 [1, 2]. Este es el principal receptor que utilisa la PTH para ejercer sus principales acciones biolÛgicas, es decir la estimulaciÛn de la resorciÛn Ûsea y del remodelaje Ûseo, la reabsorciÛn tubular de calcio, la eliminaciÛn renal de fÛsforo y la sÌntesis de calcitriol [3]. La fisiologÌa simplista de esos aÒos comenzÛ a complicarse a partir del aÒo 1987 cuando un groupo de investigadores de la universidad de Yale identificÛ el PTHrP, una proteina semejante a la PTH, resposable de los episodios hipercalcÈmicos observados en ciertos enfermos cancerosos [4, 5]. El efecto hipercalcÈmico es debido a que el PTHrP tambiÈn se fija y activa el mismo PTH-R1 en el riÒÛn y el hueso [6]. Adem·s, recientement se ha sugerido que el PTHrP es mucho mas importante que la PTH en el desarrollo Ûseo embrionario [7].
Un tercer miembro de la familia de la PTH y del PTHrP existe desde hace poco, el TIP-39 (tuberoinfundibular peptide), es un neuropÈptido el cual activa otro receptor de la PTH, el PTH-R2 [8, 9]. La expresiÛn de este ˙ltimo predomina en el p·ncreas y en el hipot·lamo. Por el momento, las funciones fisiolÛgicas del TIP-39 no se conocen con precisiÛn, pero se le atribuye una funciÛn de inhibidor natural de la PTH, sobretodo al fragmento TIP (7-39) el cual se fija al PTH-R1 sin activarlo y bloquea la acciÛn de la PTH.
La existencia del PTH-R2 y son activaciÛn al ligarse con el TIP-39 podrÌa explicar el doble effecto, estimulador e inhibidor, de la PTH sobre la proliferaciÛn celular. De hecho, el TIP-39 inhibe la proliferaciÛn de las cÈlulas HEK293 transfectadas con el PTH-R2 [10]. Adem·s, la administracion intracerebroventricular de TIP-39 reduce significantemente la concentraciÛn plasm·tica de vasopresina (ADH) en animales deshidratados, lo que sugiere un effecto central de esta molÈcula en la osmoregulaciÛn y baroregulaciÛn de la secreciÛn de la ADH [11].
TambiÈn, se ha propuesto que el TIP-39 podrÌa ser un neuromodulador de la sensibilidad al dolor [12]. Finalemente, los ARN mensajeros del TIP-39 y del PTH-R2 se han encontrado en la paredes de los vasos sanguÌneos renales, donde el TIP-39, como otros pÈptidos que se ligan al PTH-R2, tienen la facultad de vasodilatar esos vasos independientemente del PTH-R1. Esos resultados sugieren que TIP-39 y PTHrP podrÌan simultaneamente coordinar la regulaciÛn de la hemodin·mica renal [13].
Hay evidencias en favor de la existencia de otros receptores de la PTH, un tipo tres (PTH-R3), probablemente especÌfico del PTHrP en los queratinocitos, en el riÒÛn y en la placenta [7], y un tipo cuatro (PTH-R4) especÌfico para los fragmentos C terminales de la PTH [14].
El PTH-R1 ha sido el m·s estudiado de los tres receptores hasta ahora identificados. Se trata de una molÈcula de la familia de los receptores ligados a las proteinas G, los cuales tienen como caracterÌsticas estructurales principales, siete regiones en forma de hÈlice, las cuales atraviesan la membrana celular, una regiÛn extracelular donde vienen a fijarse los agonistas y una regiÛn intracelular que sirve para transmitir el mensaje por medio de las proteÌnas G.
Las dos molÈculas, PTH y PTHrP se ligan con gran afinidad al PTH-R1 y estimulan un sistema de transducciÛn que implica la activaciÛn de diferentes proteÌnas G, la fosfolipasa C y la adenilciclasa [2, 15]. El uso diferencial de esas vÌas de transducciÛn depende del asamblaje del PTH-R1 con la proteÌna NHERF (sodium-hydrogen exchanger regulatory factor). En caso de la activaciÛn de la vÌa fosfolipasa, la proteÌna NHERF es la ˙nica responsable de l'uniÛn del PTH-R1 con la fosfolipasa C beta, lo que quiere decir que la presencia o la ausencia de expresiÛn de NHERF determina en parte las acciones tisulares especÌficas de la PTH y del PTHrP [16].
El ARN mensajero y la proteÌna del PTH-R1 son casÌ ubicuos, pero su expresiÛn principal se encuentra en el riÒÛn, el hÌgado, el hueso, el pulmÛn, los testÌculos y la piel [17, 18]. En el riÒÛn, el ARN mensajero se localiza en la corteza y en la zona externa de la medular externa, no observ·ndose expressiÛn en la mÈdula interna. En las zonas positivas, las cÈlulas con PTH-R1 son los podocitos glomerulares, las cÈlulas del t˙bulo contorneado proximal y las del t˙bulo contorneado distal [19]. En el hueso, los estudios de hibridaciÛn in situ, han mostrado que el PTH-R1 es ˙nicamente presente en los condrocitos pre-hipertrÛficos del cartÌlago de crecimiento. La PTH como el PTHrP regulan en ese sitio la diferenciaciÛn y la apoptosis celular y el desarrollo normal del hueso endocondral en compaÒÌa de otras proteÌnas como el Ihh (indian hedgehog) [20-23].
Enfermedades determinadas genÈticamente
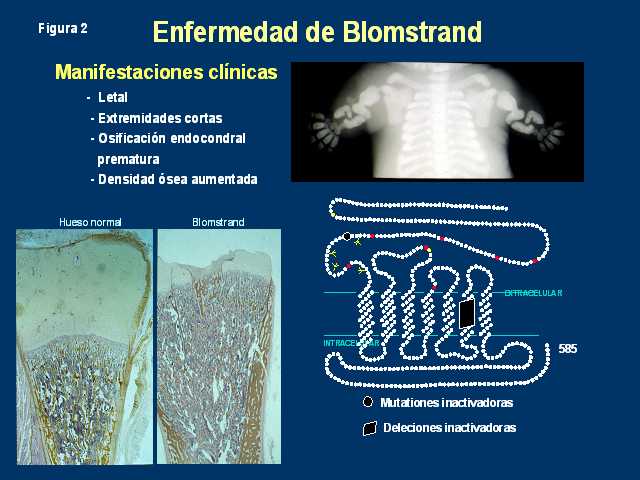
Varias mutaciones en el PTH-R1 han permitido aclarar el origen de ciertas enfermedades genÈticas [24]. Las mutaciones activadoras del PTH-R1 provocan la condrodisplasia metafisaria de tipo Jansen (fig. 1) [25] y las mutaciones inactivadoras son responsables de otra condrodisplasia rara, la enfermedad de Blomstrand (fig. 2) [26] y de ciertas encondromatosis [27].
La condrodisplasia metafisaria de Jansen es un enanismo con deformaciones esquelÈticas y osteopÛrosis. BiolÛgicamente, los enfermos presentan signos de hiperparatiroidismo con hipercalcemia, hipofÛsforemia, hipercalciuria y paradÛjicamente una concentraciÛn de PTH casi indetectable en el suero. Las mutaciones ocurren generalmente en las regiones trans-membranales y activan constitutivamente y continuamente el PTH-R1. Esta activaciÛn constante se manifiesta en un retardo de maduraciÛn de los condrocitos pre-hipertrÛficos, una falta de apoptosis y la persistencia de cÈlulas inmaduras dentro del hueso ya formado o mineralizado, dando asÌ lugar a las deformaciones Ûseas y la corta estatura.
El mismo escenario se reproduce en los animales donde se superexpresa el PTHrP especÌficamente en los condrocitos, gracias al promotor del gen del col·geno de tipo II, y cuando se introduce el PTH-R1 mutado de la enfermedad de Jansen el cual est· constantemente activado. En los dos casos se retarda la diferenciaciÛn de los condrocitos y la mineralizaciÛn Ûsea, se suspende el fenÛmeno de apoptosis y se observan islotes de condrocitos inmaduros dentro del hueso mineralizado [25, 28].
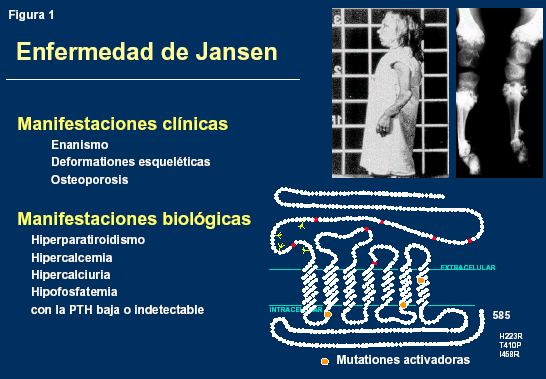
Al contrario, en la enfermedad de Blomstrand y en la encondromatosis, las mutaciones ocurren en el sexto dominio trans-membrana o en el sector extracelular e inactivan completamente el PTH-R1 [24, 29]. Los fetos mueren in ˙tero, osificados y probablemente de insuficiencia cardÌaca, puesto que en el modelo animal de knock-out del PTH-R1, los cardiomiocitos sufren una masiva y abrupta apoptosis entre los dias 11 y 12 de la vida embrionaria [30].
De hecho, esas mutaciones aceleran el proceso de maduraciÛn de los condrocitos pre-hipertrÛficos y el proceso de osificaciÛn endocondral. Los modelos animales han confirmado tambiÈn esos cuadros clÌnicos. AsÌ los ratones knock-out para los genes del PTH-R1 o de la proteÌna PTH-rP muestran el mismo fenotipo Ûseo que los enfermos con la condrodisplasia de Blomstrand. Lo que significa que en ausencia de PTHrP o del PTH-R1 los condrocitos no pueden proliferar normalmente e impident el crecimiento Ûseo longitudinal normal. Los condrocitos se diferencian mas r·pido, comienzan a producir los componentes de la matriz extracelular precozmente, facilitan la mineralizaciÛn Ûsea, y mueren de apoptosis prematura.
Adem·s, es posible que algunos polimorfismos en el gen PTH-R1 puedan determinar la sensibilidad del riÒÛn y del hueso a la acciÛn catabÛlica o anabÛlica de la PTH [31, 32].
Enfermedades adquiridas
Varias situaciones clÌnicas se associan con una alteraciÛn funcional del PTH-R1, altÈraciÛn que puede ser secundaria a la disminuciÛn de la expresiÛn del ARNm/proteÌna o a un defecto en las vÌas de transducciÛn del seÒal intracelular. Asi, la expresiÛn del ARNm del PTH-R1 y de su proteÌna es regulada, por lo menos en los principales Ûrganos sensibles a la acciÛn de la PTH, por numerosos factores.
Por un lado, las endotoxinas, la interleuquina-2, la dexametasona, la T3, el FGF-2 (fibroblast growth factor-2) y el (transforming growth factor-beta) estimulan positivamente la expressiÛn del PTH-R1 [33, 34]. El hecho que la dexametasona lo regule de manera positiva en las celulas Ûseas es uno de los mecanismos por los cuales el tratamiento con esteroides de enfermedades immunolÛgicas y de transplantes de Ûrganos se complica con perdida Ûsea. Igualmente, en los animales con dÈficit en vitamina D y sin insuficiencia renal, es com˙n encontrar un PTH-R1 aumentado en el hueso y en el riÒÛn [35].
Por otro lado, la PTH, el PTHrP, la angiotensina-II, el IGF-1 (insulin growth factor-1), la PGE2 (prostaglandin E2), la vitamina D, la insufficiencia renal aguda (IRA) y crÛnica disminuyen la expresiÛn del PTH-R1.
En la insuficiencia renal aguda, Soifer et cols., demostraron que el PTH-R1 disminuye en las cÈlulas del t˙bulo contorneado prÛximal algunas horas despuÈs de una isquemia provocada, probablemente como consecuencia de un aumento en la producciÛn de PTHrP. Lo cual hace pensar que el sistema PTH/PTHrP y su receptor podrÌan estar implicados el el proceso de reparaciÛn o agravaciÛn de la IRA [36]. El mismo sistema, actuando de manera paracrina o autocrina, y donde un exceso de producciÛn de PTHrP provoca une disminuciÛn del PTH-R1 en las cÈlulas vasculares, ha sido implicado en la fisiopatologÌa de la hipertensiÛn arterial en las ratas SHR (spontaneously hypertensive rats) [37]. Igualemente, el PTHrP y su receptor parecen jugar un papel pro-inflammatorio en las lesiones ateromatosas y en la inestabilidad de las placas atheromatosas en las arterias carÛtidas [38]. En la piel, el PTHrP es tambiÈn un potente inhibidor de la proliferaciÛn de cÈlulas escamosas. Por esta razon, ciertos estudios recientes sugieren la aplicacion local de PTHrP como tratamiento de la psoriasis [39].
De manera similar, en caso de insuficiencia renal crÛnica, el PTH-R1 diminuye progresivamente en el riÒÛn paralelamente con la disminuciÛn de la filtraciÛn glomerular [40, 41]. En el hueso, la disminuciÛn del PTH-R1 se asocia con una reducciÛn de la altura de la zona ocupada por los condrocitos pre-hipertrÛficos, como lo han demostrado los estudios de hibridaciÛn in situ [42]. Estas alteraciones son en parte responsables de la resistencia del hueso a la acciÛn hipercalcÈmica de la PTH y del retardo de crecimiento longitudinal en los niÒos urÈmicos [40-43].
El nivel de expresiÛn del PTH-R1 juega tambiÈn un papel importante en la instauraciÛn de la enfermed Ûsea adin·mica (ABD) observada en ciertos enfermos urÈmicos. En un trabajo reciente, Picton et cols. demuestran, en un grupo de 13 enfermos dializados, une disminuciÛn de mas de 70% de la expresiÛn del PTH-R1 en las cÈlulas Ûseas. Esta disminuciÛn se observa en todos los tipos de osteopatÌas urÈmicas, pero es mucho m·s importante en el hueso de los enfermos con una ABD [44].
En caso de acidosis metabÛlica la repuesta a la acciÛn de la PTH aumenta y se acompaÒa de una hiperresorpciÛn y perdida Ûsea. Ese fenÛmeno es debido a una aumentaciÛn de la expresiÛn del PTH-R1 en los osteoblastos [45].
En cuanto a los receptores de los fragmentos C-terminales de la PTH. El equipo cadaniense de d'Amour mostrÛ recientemente que en los animales tiroparatiroidectomizados, el efecto hipercalcÈmico de la PTH (1-84) es bloqueado completamente cuando se administra conjuntamente con una concentraciÛn equimolar de PTH (7-84) ou con una mezcla de fragmentos C-terminales de la PTH [46]. Igualmente, Bringhurst et cols. Demuestran, en un modelo de celulas osteobl·sticas de animales knock-out para el gen del PTH-R1, que la fijaciÛn del fragmento (19-84) de la PTH no puede desplazarse con cantidades crecientes del fragmento activo (1-34) de la PTH, lo que sugiere la existencia de un receptor especÌfico de ese fragmento C-terminal de la PTH en las cÈlulas Ûseas [14].
ConclusiÛn
Las implicaciones biolÛgicas de la identificacion y el clonaje de los receptores de la familia PTH son innumerables (tabla 1). La distribuciÛn casi general y ubicua del PTHrP y del PTH-R1 asÌ como de los diferentes fragmentos de esas molÈculas, nos invitan a crÈer en la existencia de otros receptores y que la acciÛn de esas molÈculas sobre el sistema mineral y Ûseo es mucho m·s compleja que lo que pens·bamos.
IMPLICACIONES BIOLOGICAS DEL RECEPTOR PTH-R1
Mutaciones genÈticas activadoras del PTH-R1
- Condrodisplasia de Jansen
|
Mutationes genÈticas inactivadoras del PTH-R1
- Condrodisplasia de Blomstrand
- Encondromatosis (Enfermedad de Ollier y Maffucci)
|
| Polimorfismos genÈticos determinando la sensibilidad renal y Ûsea a la PTH |
AlteraciÛn en la regulaciÛn del PTH-R1
- Corticoesteroides
- Insuficiencia renal crÛnica
- Insuficiencia renal aguda
- Enfermedad Ûsea adin·mica
- HipertensiÛn arterial espont·nea en ratas (SHR)
|
AcciÛn de la PTH, el PTHrP o el TIP-39 sobre otros receptores de la PTH
- Receptor especÌfico de los fragmentos C-terminales de la PTH
- Tratamiento tÛpico de la psoriasis con PTHrP
|
BibliografÌa
1. J¸ppner H, Abou-Samra A, Freeman M, et al.: A G protein-linked receptor for parathyroid hormone and parathyroid hormone-related peptide. Science 1991;254:1024-6
2. Abou-Samra A, J¸ppner H, Force T, et al.: Expression cloning of a common receptor for parathyroid hormone and parathyroid hormone-related peptide from rat osteoblast-like cells: a single receptor stimulates intracellular accumulation of both cAMP and inositol trisphosphates and increases intracellular free calcium. Proc Natl Acad Sci USA 1992;89:2732-6
3. UreÒa P, Abou-Samra A, J¸ppner H, et al.: Mode d'action de l'hormone parathyroÔdienne (PTH) et du peptide apparantÈ ý la PTH (PTHrP) au niveau des organes cibles. Annales d'Endocrinologie 1994;55:133-41
4. Strewler G: The physiology of parathyroid hormone-related protein. N Engl J Med 2000;342:177-85
5. Burtis W, Wu T, Bunch C, et al.: Identification of a novel 17,000-Dalton parathyroid hormone-like adenylate cyclase-stimulating protein from a tumor associated with humoral hypercalcemia of malignancy. J Biol Chem 1987;262:7151-6
6. Orloff J, Wu T, Stewart A: Parathyroid hormone-like proteins: biochemical responses and receptor interactions. Endocrine Reviews 1989;10:476-95
7. Orloff J, Kats Y, UreÒa P, et al.: Further evidence for a novel receptor for amino-terminal parathyroid hormone-related protein on keratinocytes and squamous carcinoma cell lines. Endocrinology 1995;136:3016-23
8. Usdin T, Gruber C, Bonner T: Identification and functional expression of a receptor selectively recognizing parathyroid hormone, the PTH2 receptor. J Biol Chem 1995;270:15455-8
9. Usdin T: Evidence for a parathyroid hormone-2 receptor selective ligand in the hypothalamus. Endocrinology 1997;138:831-4
10. Misiano P, Scott BB, Scheideler MA, Garnier M: PTH2 receptor-mediated inhibitory effect of parathyroid hormone and TIP39 on cell proliferation. Eur J Pharmacol 2003;468:159-66
11. Sugimura Y, Murase T, Ishizaki S, et al.: Centrally administered tuberoinfundibular peptide of 39 residues inhibits arginine vasopressin release in conscious rats. Endocrinology 2003;144:2791-6
12. Usdin TB, Dobolyi A, Ueda H, Palkovits M: Emerging functions for tuberoinfundibular peptide of 39 residues. Trends Endocrinol Metab 2003;14:14-9
13. Eichinger A, Fiaschi-Taesch N, Massfelder T, et al.: Transcript expression of the tuberoinfundibular peptide (TIP)39/PTH2 receptor system and non-PTH1 receptor-mediated tonic effects of TIP39 and other PTH2 receptor ligands in renal vessels. Endocrinology 2002;143:3036-43
14. Divieti P, John M, J¸ppner H, Bringhurst F: Human PTH-(7-84) inhibits bone resorption in vitro via actions independent of the type I PTH/PTHrP receptor. Endocrinology 2002;143:171-6
15. J¸ppner H: Functional properties of the PTH/PTHrP receptor. Bone 1995;17:39S-42S
16. Mahon MJ, Donowitz M, Yun CC, Segre GV: Na(+)/H(+ ) exchanger regulatory factor 2 directs parathyroid hormone 1 receptor signalling. Nature 2002;417:858-61
17. UreÒa P, Kong X, Abou-Samra A, et al.: Parathyroid hormone (PTH) /PTH-related peptide (PTHrP) receptor mRNA are widely distributed in rat tissues. Endocrinology 1993;133:617-23
18. UreÒa P, Lee K, Weaver D, et al.: PTH/PTHrP receptor mRNA expression as assessed by Northern blot and in situ hybridization. J Bone Miner Res 1992;7:S118
19. Lee K, Brown D, UreÒa P, et al.: Localization of parathyroid hormone/parathyroid hormone-related peptide receptor mRNA in kidney. Am J Physiol 1996;270:F186-F91
20. Vortkamp A, Lee K, Lanske B, et al.: Regulation of rate of cartilage differentiation by indian hedgehog and PTH-related protein. Science 1996;273:613-21
21. Lanske B, Karaplis A, Lee K, et al.: PTH/PTHrP receptor in early development and indian hedgehog-regulated bone growth. Science 1996;273:663-6
22. Lee K, Deeds J, Bond A, et al.: In situ localization of PTH/PTHrP receptor mRNA in the bone of fetal and young rats. Bone 1993;14:341-5
23. Lee K, Deeds J, Segre G: Expression of parathyroid hormone-related peptide and its receptor messenger ribonucleic acids during fetal development of rats. Endocrinology 1995;136:453-63
24. Silve C: RÈcepteur PTH/PTHrP et maladies gÈnÈtiques. MÈdecine/Sciences 1999;15:1310-2
25. Schipani E, Kruse K, J¸ppner H: A constitutively active mutant PTH-PTHrP receptor in Jansen-type metaphyseal chondrodysplasia. Science 1995;268:98-100
26. Zhang P, Jobert A, Couvineau A, Silve C: A homozygous inactivating mutation in the parathyroid hormone/parathyroid hormone-related peptide receptor causing Blomstrand chondrodysplasia. J Clin Endocrinol Metab 1998;83:3365-8
27. Hopyan S, Gokgoz N, Poon R, et al.: A mutant PTH/PTHrP type I receptor in enchondromatosis. Nature Genetics 2002;30:306-10
28. Weir E, Philbrick W, Amling M, et al.: Targeted overexpression of parathyroid hormone-related peptide in chondrocytes causes chondrodysplasia and delayed endochondral bone formation. Proc Natl Acad Sci 1996;93:10240-5
29. J¸ppner H, Gardella T, Lee C, et al.: Mapping of the ligand binding domain by using mutant PTH/PTHrP receptors. J Bone Miner Res 1993;7:S93
30. Qian J, Colbert MC, Witte D, et al.: Midgestational lethality in mice lacking the parathyroid hormone (PTH)/PTH-related peptide receptor is associated with abrupt cardiomyocyte death. Endocrinology 2003;144:1053-61
31. Heishi M, Tazawa H, Matsuo T, et al.: A novel Van91 I polymorphism in the 1st intron of the parathyroid hormone (PTH)/PTH-related peptide (PTHrP) receptor gene and its effect on the urinary cAMP response to PTH. Biological and Pharmaceutical Bulletin 2000;23:386-9
32. Minagawa M, Yasuda T, Watanabe T, et al.: Association between AAAG repeat polymorphism in the P3 promoter of the human parathyroid hormone (PTH)/PTH-related peptide receptor gene and adult height, urinary pyridinoline excretion, and promoter activity. J Clin Endocrinol Metab 2002;87:1791-6
33. Mannstadt M, Dr¸eke T: RÈcepteurs de l'hormone parathyroÔdienne: du clonage aux implications physiologiques, physiopathologiques et cliniques. NÈphrologie 1997;18:5-10
34. UreÒa P, Iida-Klein A, Kong X, et al.: Regulation of the parathyroid hormone (PTH)/PTH-related peptide receptor mRNA by glucocorticoids and PTH in ROS 17/2.8 cells and OK cells. Endocrinology 1994;134:451-6
35. Turner G, Coureau C, RiszkRabin M, et al.: Parathyroid hormone (PTH)/PTH-related protein receptor messenger ribonucleic acid expression and PTH response in a rat model of secondary hyperparathyroidism associated with vitamin D deficiency. Endocrinology 1995;136:3751-8
36. Soifer N, VanWhy S, Ganz M, et al.: Expression of parathyroid hormone-related protein in the rat glomerulus and tubule during recovery from renal ischemia. J Clin Invest 1993;92:2850-7
37. Massfelder T, Taesch N, Fritsch S, et al.: Type 1 parathyroid hormone receptor expression level modulates renal tone and plasma renin activity in spontaneously hypertensive rat. J Am Soc Nephrol 2002;13:639-48
38. Martin-Ventura JL, Ortego M, Esbrit P, et al.: Possible role of parathyroid hormone-related protein as a proinflammatory cytokine in atherosclerosis. Stroke 2003;34:1783-9
39. Holick MF, Chimeh FN, Ray S: Topical PTH (1-34) is a novel, safe and effective treatment for psoriasis: a randomized self-controlled trial and an open trial. Br J Dermatol 2003;149:370-6
40. UreÒa P, Mannstadt M, Hruby M, et al.: Down-regulation of the PTH/PTHrP receptor in uremia. J Bone Min Metab 1994;12:S87-S90
41. UreÒa P, Kubrusly M, Mannstadt M, et al.: The renal PTH/PTHrP receptor is downregulated in rats with chronic renal failure. Kidney Int 1994;45:605-11
42. UreÒa P, Ferreira A, Morieux C, Dr¸eke T, deVernejoul M: PTH/PTHrP receptor mRNA is down-regulated in epiphyseal cartilage growth plate of uraemic rats. Nephrol Dial Transplant 1996;11:2008-16
43. UreÒa P, Mannstadt M, Hruby M, et al.: Parathyroidectomy does not prevent the renal PTH/PTHrP receptor down-regulation in uremic rats. Kidney Int1995;47:1797-805
44. Picton M, Moore P, Mawer E, et al.: Down-regulation of human osteoblast PTH/PTHrP receptor mRNA in end-stage renal failure. Kidney Int 2000;58:1440-9
45. Disthabanchong S, Martin KJ, McConkey CL, Gonzalez EA: Metabolic acidosis up-regulates PTH/PTHrP receptors in UMR 106-01 osteoblast-like cells. Kidney Int 2002;62:1171-7
46. Nguyen-Yamamoto L, Rousseau L, Brossard J, Lepage R, D'Amour P: Synthetic carboxy-terminal fragments of parathyroid hormone (PTH) decrease ionized calcium concentration in rats by acting on a receptor different from the PTH/PTHrP-related peptide receptor. Endocrinology 2001;142:1386-92
