Introducción
La importancia de las apolipoproteínas en el sistema nervioso central (S.N.C.) surgió con la asociación del alelo E4 de la apolipoproteína E (apo E) con un tipo familiar de enfermedad de Alzheimer (EA). Estudios posteriores han demostrado que la apo E y J están relacionadas con la formación de las placas seniles y los depósitos amiloideos de los vasos. Entre las apolipoproteínas sintetizadas en el S.N.C. destacan la apo E, J, C-1 y D. Se cree que estas apolipoproteínas (al menos la apo D y E) están posiblemente relacionadas con el transporte, almacenamiento y redistribución de los lípidos cerebrales. La apo D es una glicoproteína plasmática que se localiza en el tejido nervioso humano (células de glía, endoteliales, perivasculares y neuronas).En el presente trabajo hemos estudiado la localización y presencia de apo D en las lesiones vasculares de la angiopatía cerebral amiloidea (A.C.A.).
Material y Métodos
Para la realización del estudio hemos utilizado un anticuerpo policlonal para la apolipoproteína D (cedido por el Dr. C. López Otín) y otro para la apolipoproteína E (Chemicon), así como una detección histoquímica para amiloide. El material utilizado procede de necropsias realizadas por el Servicio de Anatomía Patológica del Hospital Central de Asturias.
Resultados
Los resultados obtenidos muestran la presencia de apo D en gran parte de las lesiones vasculares de la ACA. La apo D aparece tanto en los vasos piales, como en los corticales y subcorticales, siempre relacionada con el tipo de lesiones que presentaba el sujeto. En los capilares y vasos de pequeño calibre, la apo D aparece en las células endoteliales y perivasculares. En vasos de mayor calibre la inmunorreactividad es siempre adventicial con poca o ninguna infiltración en las capas endotelial y media.
Discusión
La función de la apo D en los vasos sangíneos en la ACA podría estar relacionada con la formación del depósito amiloide o por otra parte estar comprometida en el transporte de colesterol (y otras moléculas hidrofóbicas) en la zona debido a la deficiente función de la apo E y a la lesión de la barrera hematoencefálica.
PALABRAS CLAVE: amiloide, Rojo Congo, apolipoproteína D, apolipoproteína E, inmunohistoquímica.
La angiopatía cerebral amiloidea (ACA) se define como la deposición de material congófilo (fibrillas insolubles de amiloide) en vasos sanguíneos corticales y meníngeos. Es una patología que ocurre frecuentemente en ancianos, independientemente de que estos presenten demencia o no. La presencia de amiloide en los vasos cerebrales es un riesgo, en fases avanzadas de ACA, para la ruptura vascular y la hemorragia cerebral (1).
El análisis del amiloide ha demostrado que se trata de un péptido derivado de una proteína transmembrana denominada proteína precursora de amiloide (PPA). Esta proteína es anormalmente procesada dando lugar a péptidos de 30-40 (Aß40) o de 42-43 (Aß42) residuos aminoacídicos. Dichos péptidos se agregan para formar el depósito de amiloide.
La formación y progresión del deposito de amiloide ha sido bien estudiada en las placas seniles, lesiones características de la enfermedad de Alzheimer (EA). En estas estructuras existen ambos péptidos, siendo más abundante el Aß42. La deposición se iniciaría por la agregación de péptidos Aß solubles alrededor de un núcleo previamente formado. Dado que el Aß42 es relativamente más insoluble que el Aß40, podría ser que cierta cantidad de éste formase las primeras fibrillas insolubles que crecerían rápidamente por adición de Aß40. (2)
En la ACA ha sido descrita la preponderancia del péptido Aß40 sobre el Aß42, con lo que el proceso y mecanismo de deposición del amiloide en los vasos podría ser diferente al que ocurre en las placas seniles. Según algunos autores el proceso de formación del amiloide se iniciaría con la deposición de un núcleo vascular inicial de Aß42, proceso claramente independiente del alelo e4 de la apo E y que tiene tendencia a ocurrir en segmentos específicos de los vasos. Posteriormente existiría un crecimiento de los depósitos iniciales reemplazando la pared vascular, proceso que se efectuaría a partir de Aß40 y que se ha asociado con la presencia del alelo e4. Finalmente la ruptura vascular y posterior hemorragia parecen estar asociadas con el alelo e2 de la apo E (2,3).
Parece claro, hoy en día, que la nucleación del péptido Aß42 no es el único factor que determina la formación del amiloide y que existen factores externos que influyen en la unión progresiva del Aß40 al núcleo inicial. Estos factores son poco conocidos, pero algunas moléculas han demostrado que pueden jugar un papel importante como las apolipoproteinas, proteínas asociadas a la inflamación, proteoglicanos, proteasas, etc. (4).
Entre las apolipoproteinas que son sintetizadas en el sistema nervioso central (S.N.C.), destacan la apo E, J, C-1 y D. La apo E se ha involucrado directamente en la formación del depósito de amiloide, aunque su papel no ha sido todavía establecido. La apo E podría unirse al Aß favoreciendo su deposición tisular. Se ha sugerido que otras apolipoproteinas podrían tener efectos opuestos, así, la apo J podría tener influencia negativa sobre la fibrilogénesis del amiloide (5).
Nuestro interés por el estudio de la apo D surge del hecho que junto a la apo E comparte algunas propiedades como su síntesis a nivel del SNC, la capacidad de transportar pequeños ligandos hidrofóbicos y el incremento de su expresión durante la regeneración del sistema nervioso periférico. Su presencia ha sido demostrada en astrocitos, células piales, oligodendrocitos, células perivasculares y algunas poblaciones neuronales (5,6). Sin embargo en muchos casos se desconoce si esta presencia es debida a la síntesis o a la captación de apo D por parte de estas células.
En un trabajo anterior hemos demostrado la presencia de apo D en placas seniles (difusas y neuríticas) tanto de ancianos normales como aquejados de EA.(7). En el presente trabajo se ha estudiado la presencia y localización de apo D en las lesiones vasculares características de la ACA y también se ha realizado una comparación con la apo E.
Doce cerebros con ACA diagnosticados "postmortem" fueron seleccionados del Servicio de Anatomía Patológica del Hospital Central de Asturias y fueron separadas de acuerdo con el grado de ACA (1). Bloques de la corteza cerebral fueron fijados con formol, embebidos en parafina y seccionados a 10 µm de grosor. En todas las secciones se realizó una detección histoquímica previa para amiloide (según una técnica desarrollada en nuestro laboratorio y que permite una detección rápida, constante y compatible con las técnicas inmunocitoquímicas (8).
Posteriormente se realizaron detecciones inmunohistoquímicas para apo D (anticuerpo cedido por Dr. Carlos López-Otín) o para apo E (Chemicon, AB-947). Las secciones, desparafinadas y rehidratadas, fueron tratadas con suero de bloqueo en PBS. Las secciones fueron incubadas durante toda la noche con los anticuerpos primarios (diluidos, 1:1000 para la apo D, 1:500 para la apo E) y posteriormente la immunoreactividad fue detectada mediante el kit universal "Quick" de Vector. La actividad peroxidasa fue visualizada mediante la incubación con D.A.B. y la actividad fosfatasa mediante la incubación con "Vector blue substrate" (Vector). Las secciones fueron fotografiadas en un microscopio" Nikon Labophot" con una cámara digital Nikon (Colpix 950).
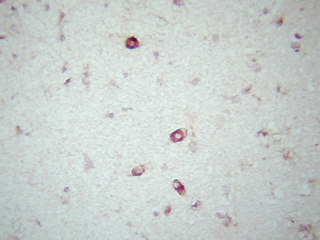
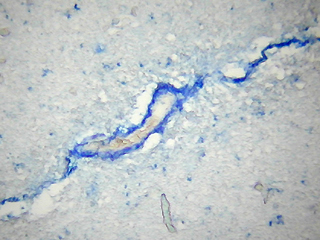
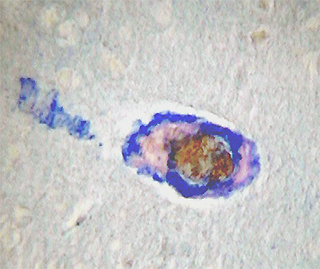
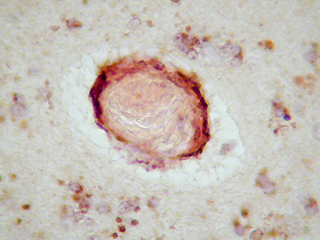
En el estudio inmunohistoquímico para la apo D de la corteza cerebral de individuos con varios grados de ACA, hemos encontrado que en muy baja proporción existe colocalización de apo D con el amiloide vascular congófilo. En los capilares perforantes y vasos de pequeño calibre con paredes engrosadas no congófilas, la apo D aparece localizada en las células endoteliales y perivasculares (Fig. 1). En los vasos piales y corticales de mayor calibre la apo D puede estar presente o no (Fig. 2), pero cuando lo hace su presencia parece inversamente relacionada con la cantidad de depósito amiloideo. De manera que, el vaso presenta una fuerte inmunorreactividad para la apo D cuando la cantidad de amiloide es escasa o nula y las paredes apenas están afectadas (Fig. 3). Esta se presenta siempre en la adventicia o rodeando el pequeño depósito de amiloide (Fig. 4). Por otra parte, cuando el depósito congófilo es grande y bien desarrollado alrededor del diámetro vascular como ocurre en los casos de ACA media y severa, la inmunorreactividad para la apo D es pequeña o inexistente (Fig. 5).
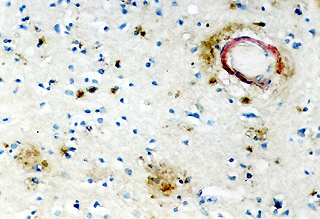
Comparando estos hallazgos, con los obtenidos para la apo E, hemos comprobado que ambas apolipoproteinas se comportan de forma distinta. Así, las lesiones vasculares congófilas son fuertemente inmunorreactivas para la apo E, independientemente de la cantidad de amiloide y del tipo o situación del vaso. La inmunotinción para la apo E puede ser tan fuerte que llega a enmascarar la presencia de amiloide (Fig. 6). Incluso los capilares perforantes no congófilos lesionados presentaban en ocasiones inmunomarcaje para la apo E.
El estudio inmunohistoquímico de la corteza cerebral de pacientes con ACA diagnosticada "postmortem" mediante una modificación del Rojo Congo (8) revela la presencia de la apo D en las lesiones vasculares preamiloideas así como también en algunas con depósito congófilo.
Algunas apolipoproteinas se sabe que están íntimamente asociadas con la fibrilogénesis del amiloide. La apo E y otros factores pueden acelerar la formación de las fibrillas de amiloide alrededor de los vasos o en las placas seniles. Sin embargo, se ha demostrado que la apo J (y también quizás otros factores), tendrían un efecto negativo en la fibrilogénesis (9). También la A1 se ha encontrado en placas seniles y puede formar fibrillas congófilas (10). Es posible que la formación de amiloide esté determinada por un balance entre los factores promotores y los preventivos de la fibrilogénesis (2). La presencia de apo D en los depósitos preamiloideos y amiloideos vasculares de la ACA, junto nuestra descripción en un trabajo anterior de la presencia de apo D en las placas seniles (7), parece señalar algún tipo de papel para la apo D en la amilogénesis. Sin embargo no ha sido publicado hasta ahora apo D en la formación de fibrillas del ß-amiloide.
En algunas neuropatologías (que incluyen la EA) la cantidad de ARNm de la apo D en hipocampo y de apo D en líquido cefalorraquídeo (LCR) humano está incrementada comparada con controles normales (11). Parece probable que como miembro de la familia de las lipocalinas, la apo D transporte pequeños ligandos hidrófobos esenciales, sobretodo en situaciones patológicas (11). Ha sido postulado que la apo D podría facilitar el transporte de colesterol a través de la barrera hemato-encefálica (BHE) de monos (12). Dentro de la ACA, la presencia de apo D en la pared adventicial y rodeando los depósitos congófilos, podría ser debida al aumento de los requerimientos de transporte del colesterol o por otra parte a un deficiente funcionamiento de la BHE debido a la lesión vascular.
La expresión del gen de la apo D ha sido asociada con la detención del crecimiento (13) y también como respuesta no específica al daño y muerte celular (14). Sin embargo no siempre esto es así pues en el SNP, la sobreexpresion de apo D está ligada al proceso de regeneración (15). Se podría pensar que la presencia de apo D es un buen indicativo de que se está efectuando un transporte activo de moléculas hidrofóbicas (acido araquidónico, esteroles, esteroides, etc.) que serían necesarias para la reparación del tejido nervioso después del daño celular, efectuando funciones coordinadas con la apo E. El incremento en la presencia de la apo D en las lesiones amiloideas cerebrales podría interpretarse como un mecanismo compensador ante la falta de eficacia de la apo E.
(1) Vonsattel JPG, Myers RH, Hedley-Whyte ET, Ropper AH, Bird ED, Richardson EP Jr. Cerebral amyloid angiopathy without and with cerebral hemorrhages: a comparative histological study. Ann Neurol 1991; 30: 637-649.
(2) Verbeek MM, Eikeleboom P, de Waal RMW. Differences between the pathogenesis of senile plaques and congophilic angiopathy in Alzheimer disease. J Neuropathol Exp Neurol 1997; 56 (7): 751-761.
(3) Alonzo NC, Hyman BT, Rebeck GW, Greenberg SM. Progression of cerebral amyloid angiopathy: accumulation of amyloid-ß40 in affected vessels. J Neuropathol Exp Neurol 1998; 57(4): 353-359.
(4) Dickson DW. The pathogenesis of senile plaques. J Neuropathol Exp Neurol 1997; 56(4): 321-339.
(5) Beffert U, Danik M, Krywkowski P, Ramassamy C, Berrada F, Poirier J. The neurobiology of apolipoproteins and their receptors in the CNS and Alzheimer´s disease. Brain Res Rev 1998; 27(2): 119-142.
(6) Navarro A, Tolivia J, Astudillo A, del Valle E. Pattern of apolipoprotein D immunoreactivity in human brain. Neurosci Lett 1998; 254: 17-20.
(7) Navarro A, Astudillo A, del Valle E., González del rey C, Tolivia, J. Immunohistochemical presence of apolipoprotein D in senile plaques. J Histotech 2000 (in press).
(8) Navarro A, Tolivia J, del Valle E. Congo red method for demonstrating amyloid in paraffin sections. J Histotech 1999; 22(4): 305-308.
(9) Oda T, Wals P,Osterburg HH, Jhonson SA, Pasinetti GM, Morgan TE, Rozovsky I, Stine WB, Snyder SW, Holzman TF, Kraft GA, Finch CE. Clusterin (apo J) alters the aggregation of amyloid ß-peptide (Aß 1-42) and forms slowly sedimenting Aß complexes that cause oxidative stress. Exp Neurol 1995; 136(1): 22-31.
(10) Wisniewski T, Golabek AA, kida E, Wisniewski KE, Frangione, B. Conformational mimicry in Alzheimer´s disease. Role of apolipoproteins in amyloidogenesis. Am J Pathol 1995, 147 (2): 238-244.
(11) Terrisse L, Poirier J, Bertrand P, Merched A., Visvikis S, Siest G, Milne R, Rassart E. Increased levels of apolipoprotein D in cerebrospinal fluid and hippocampus of Alzheimer´s patients. J Neurochem 1998, 71(4): 1643-1650.
(12) Smith K, Lawn R, Wilcox J. Cellular localization of apolipoprotein D and lecithin:cholesterol acyltransferase mRNA in rhesus monkey tissues by in situ hybridization. J Lipid Res 1990, 31: 995-1004.
(13) Provost PR, Marcel YL, Milne RW, Weech PK, Rassart E. Apolipoprotein D transcription occurs specifically in nonproliferating quiescent and senescent fibroblast cultures. FEBS Lett 1991, 290: 139-141.
(14) Ong WY, He Y, Suresh S, Patel SC. Differential expression of aolipoprotein D and apolipoprotein E in the kainic acid-lessioned rat hippocampus. Neurosci 1997, 79: 359-367.
(15) Spreyer P, Schaal H, Khun G, Rothe T, Unterbeck A, Cleck K, Müller HW. Regeneration-associated high level expression of apolipoprotein D mRNA in endoneurial fibroblasts of peripheral nerve. EMBO J 1990, 9: 2479-2484.