|
|
IMPLANTES
DE NERVIO CIÁTICO O DE CÉLULAS DE SCHWANN
Autores: Cid Torres M.A.; Paíno C.L.; López J.; Reimers Cerdá D.; Martín del Río R. y Herranz A.S.* Institución:
Hospital Ramón y
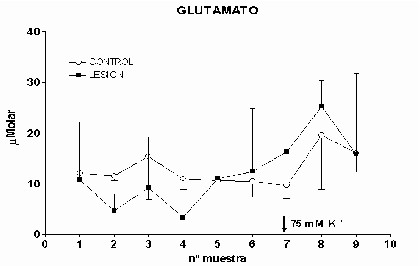
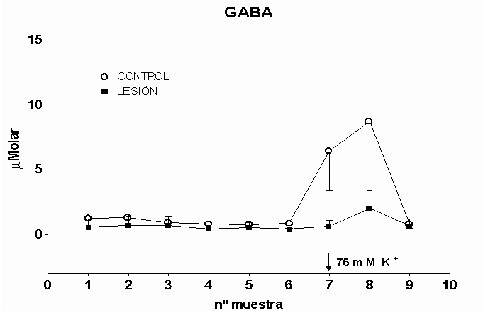
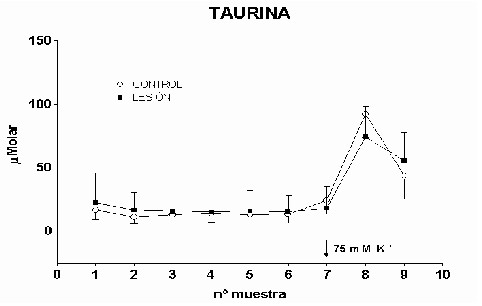
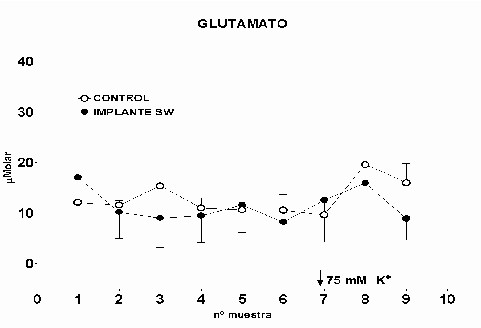
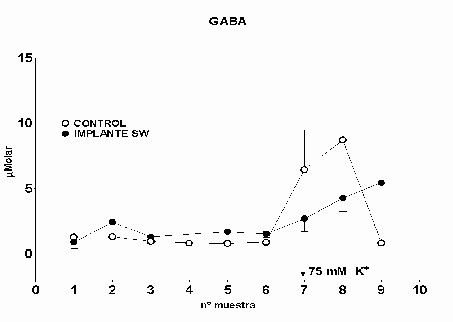
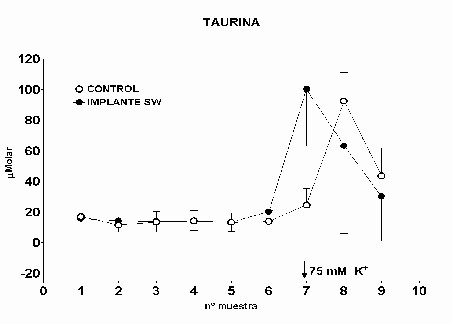
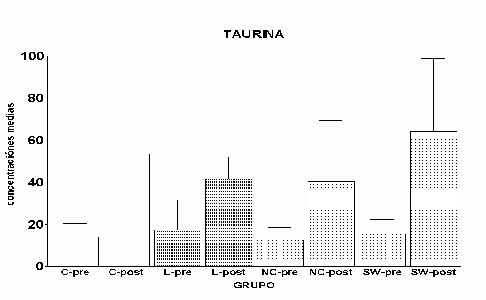
Cajal. Serv. de Neurobiología, Investigación. Email: antonio.sanchez@hrc.es Resumen Introducción y Objetivos Metodología Resultados Discusión Conclusiones Bibliografía Enviar Comentarios Los implantes de nervio ciático en corteza prefrontal lesionada provocan una recuperación en los niveles de recaptación de glutamato en el estriado ipsilateral a la lesión. Esta recuperación podría estar mediada por factores tróficos procedentes del implante de nervio ciático o de las células de Schwann presentes en él. Nuestro objetivo fue comparar los efectos de implantes de nervio ciático o de células de Schwann en animales a los que previamente se les había practicado una lesión por aspiración en la corteza prefrontal. Para ello se valoró la concentración extracelular en el estriado dorsomedial de glutamato, GABA y taurina mediante microdiálisis durante un periodo control y tras una despolarización con K+ 75 mM. En los animales lesionados se abolió la liberación de GABA inducida por K+. Los implantes de células de Schwann y de nervio ciático en la zona lesionada normalizaron los niveles de GABA tras la despolarización. Las lesiones unilaterales de la corteza prefrontal destruyen las proyecciones corticoestriatales y producen un gran número de alteraciones del comportamiento (Patrissi, G. y Stein, D. G., 1975), además de déficits en el aprendizaje de tareas que requieren alternancia (Kesslak, J. P. y cols. ,1986) o relacionadas con la orientación espacial (Dunnet, S. B. y cols. , 1987). Algunas de estas alteraciones también se inducen al lesionar el cuerpo estriado (Bjorklund, A. y cols. , 1994; Dobrossy, M. D. y Dunnett, S. B., 1997). Dichas alteraciones se recuperan espontáneamente transcurridos varios meses (Patrissi, G. y Stein, D. G., 1975, Freed, W. J. y Cannon-Spoor, E., 1989). Con el fin de normalizar dichos deficiencias, se han realizado diversos tipos de implantes en la zona lesionada, como: tejido nervioso embrionario (Dunnet y cols., 1987; Freed, W. J. y Cannon-Spoor, E., 1989), tejido periférico adulto (Collier, T. J. y Springer, J. E., 1991), y cultivos de células gliales (Isono, M. y cols. , 1992). Todos ellos han producido mejorías parciales en las deficiencias comportamentales. En animales lesionados en la corteza prefrontal Herranz y cols. (1993), valoraron los niveles de captación de glutamato en el estriado dorsomedial como medida indirecta de funcionalidad de la vía corticoestriatal. La lesión cortical producía un drástico descenso en dichos niveles. Con objeto de promover la normalización de la vía dañada se siguieron tres tipos de estrategias. En primer lugar se efectuaron implantes de corteza fetal, que no ofrecieron resultados positivos. En segundo lugar se coimplantó corteza fetal y nervio ciático adulto, el coimplante produjo una ligera recuperación y por último, estos autores implantaron injertos de nervio ciático en la zona lesionada, induciendo la normalización de los niveles de captación de glutamato. Estas observaciones parecen indicar que el papel que juega la reinervación inducida por los injertos de tejido fetal, en el restablecimiento de las funciones del sistema corticoestriatal, es menor que la inducida por los factores de tipo "trófico" que provienen de los injertos de nervio ciático. El objetivo de este trabajo fue estudiar, en animales lesionados en la corteza prefrontal, los efectos que sobre la neurotransmisión aminoacídica producían los implantes de nervio ciático y los de células de Schwann. Para ello se determinó la concentración extracelular de glutamato, GABA y taurina en el estriado dorsomedial ipsilateral a la lesión. Los animales utilizados fueron ratas hembras Sprague-Dawley, de 200 g. de peso. Grupos experimentales: Se diseñaron cuatro grupos experimentales en los que se determinó la concentración extracelular de glutamato, GABA y taurina en la región dorsomedial del estriado, ipsilateral a la lesión, mediante microdiálisis. En el primero (L) se practicó una lesión en la corteza prefrontal. El segundo grupo (C) estuvo formado por animales con una falsa lesión, sometidos únicamente al estrés quirúrgico. En ambos grupos transcurridas ocho semanas postlesión se valoraron los aminoácidos. En el tercer (NC) y cuarto grupo (SW), a las dos semanas postlesión, se les implantó en la corteza prefrontal lesionada un fragmento de nervio ciático y células de Schwann, respectivamente. En estos dos últimos experimentos los niveles extracelulares de los aminoácidos se valoraron a las seis semanas tras el implante Cirugía: La lesión en la corteza prefrontal se realizo por aspiración con una bomba de vacío, cuidando de no dañar el cuerpo calloso. Desde el bregma la lesión abarcó 3 mm anteriores y 3.5 mm posteriores, extendiéndose lateralmente 2.5 - 3 mm en la corteza parietal. Los implantes de nervio ciático se obtuvieron de un individuo adulto. Los de células de Schwann, procedieron de un cultivo de origen perinatal, siguiendo el método de Brockes y cols. (1979). Microdiálisis: El protocolo de los experimentos de microdiálisis fue como sigue: la sonda se implantó siguiendo el atlas de Patxinos y Watson, en las coordenadas: AP: – 0.8 bregma; L: 3.25 mm; y V: 3.5 mm desde la corteza. Se tomaron muestras de 15 min. , a un flujo continuo de 2.5 m l /min. . La secuencia de perfusión fue: a) periodo control o prepulso, de seis muestras, en el que se perfundió Krebs-Ringer-Bicarbonato (KRB), b) periodo de despolarización o postpulso constituido(tres muestras), durante la primera se perfundió KRB suplementado con K + 75 mM, y en las dos últimas se perfundió de nuevo con KRB. Determinación de aminoácidos: La determinación de la concentración de glutamato, GABA y taurina en las muestras obtenidas se realizó por HPLC (Herranz y cols, 1985). Los datos de cada aminoácido se analizaron mediante regresiones múltiples en dos comparaciones distintas: a) análisis individualizado, cada grupo experimental frente al control y b) análisis global, todos los grupos experimentales frente al control. Histología: Una vez realizada la microdiálisis, los cerebros se fijaron mediante perfusión intracardíaca con PBS y paraformaldehído al 4% incluyéndose en parafina posteriormente. Las secciones histológicas de la zona de interés, de 10 m m de grosor, se inmunomarcaron para proteína fibrilar ácida de glía (GFAP), y se revelaron con peroxidasa diaminobenzidina (DAB) (Hsu S.M. y cols. , 1981). Los núcleos se contrastaron con hematoxilina Harris. Grupo L: Los niveles extracelulares de glutamato fueron iguales a los del grupo C a lo largo de todo el experimento (Fig.1). El pulso despolarizante con K+ provoco en ambos grupos un aumento significativo en la concentración de glutamato respecto al periodo control (p= 0.0192). En el caso de GABA, los niveles basales durante el periodo control fueron iguales a los del grupo C. En el periodo postpulso, el grupo L no mostró cambios en GABA, por el contrario en el grupo C se produjo un aumento significativo (p= 0.0006), (Fig.2). Los niveles extracelulares de taurina fueron iguales en ambos grupos al lo largo de todo el experimento (Fig.3). El aumento de taurina inducido por K+, tanto en L cómo en C, fue muy significativo al compararlo con el periodo control (p= 0.0000), (Fig.3).
Fig. 1: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Lesión: n=3.
Fig. 2: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Lesión: n=6.
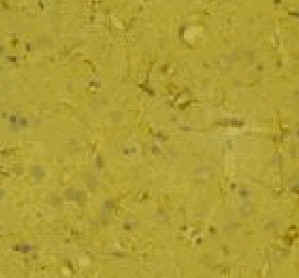
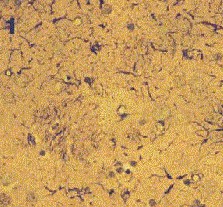
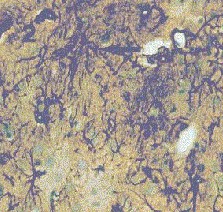
Fig. 3: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Lesión: n=8. Los resultados histológicos, mostraron un aumento en el número de astrocitos reactivos en el estriado ipsilateral a la lesión, respecto al contralateral (Fig.4)
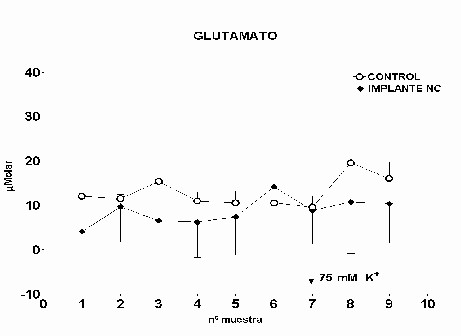
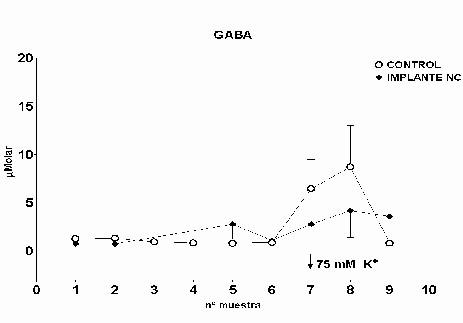
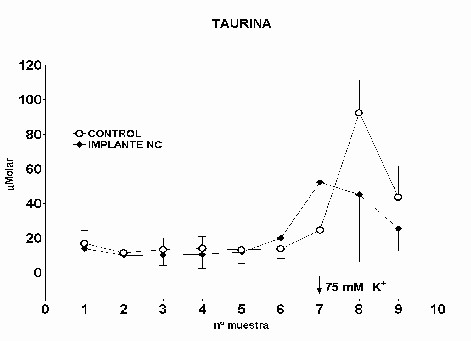
Fig.: 4: Comparación del estriado en ambos hemisferios del grupo lesionado. Inmunomarcaje para GFAP, revelado con peroxidasa-DAB. Región del estriado dorsomedial ipsi (panel derecho) y contralateral (panel izquierdo) a la lesión. Cabe destacar la reacción gliótica en panel derecho. Grupo NC: En este grupo, en el que se implanto nervio ciático, la concentración extracelular de glutamato fue significativamente menor que en el grupo C (p= 0.0163), (Fig.5). El pulso de K+ no produjo ningún cambio en los niveles extracelulares de este aminoácido. La concentración extracelular GABA no mostró diferencias entre ambos grupos a lo largo de todo el experimento. La despolarización con K+ provocó en ambos grupos un incremento en la concentración respecto a los valores basales del periodo control (p= 0.0000), (Fig.6). En el caso de taurina los niveles extracelulares fueron similares en los grupos NC y C durante todo el experimento. En el periodo de despolarización se observó un aumento significativo en ambos grupos respecto al periodo control (p= 0.0000), (Fig.7).
Fig. 5: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Implante NC: n=4.
Fig. 6: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Implante NC: n=4.
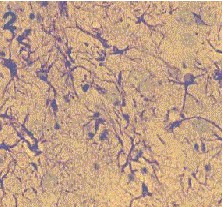
Fig. 7: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Implante NC: n=4. El inmunomarcaje revelo en el estriado ipsilateral al implante un incremento en la densidad de astrocitos reactivos, así como en el número y calibre de sus proyecciones (Fig.8)
Fig. 8: Implantes de nervio ciático. Región dorsomedial del estriado, ambos hemisferios. Inmunomarcaje para GFAP, revelado con peroxidasa-DAB. Hemisferio contralateral (1) e implantado (2). Nótese la mayor reactividad en este último Grupo SW: La concentración extracelular de glutamato fue similar durante todo el experimento en los grupos SW y C (Fig.9). La concentración extracelular de GABA durante el periodo control no mostró diferencias significativas entre ambos grupos SW y C (Fig.10). El pulso despolarizante provocó en ambos grupos un aumento muy significativo respecto al periodo control (p= 0.0000), siendo los niveles de GABA del grupo SW menores que los del grupo C (p= 0.0456). Los niveles de taurina no mostraron diferencias estadísticas entre el grupo SW y el C a lo largo de todo el experimento (Fig.11). El aumento en la liberación de taurina, durante el periodo postpulso, fue significativo en ambos grupos al compararlo con el periodo control (p= 0.0000).
Fig. 9: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Implante SW: n=6.
Fig. 10: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Implante SW: n=6.
Fig. 11: Los valores representados corresponden a la media ± desviación estándar. Control: n=3; Implante SW: n=6. El análisis inmunohistoquímico mostró que la región dorsomedial del estriado, ipsilateral al implante, presentaba un gran número de astrocitos reactivos en los que la mayoría de las proyecciones se encontraban muy engrosadas (Fig. 12)
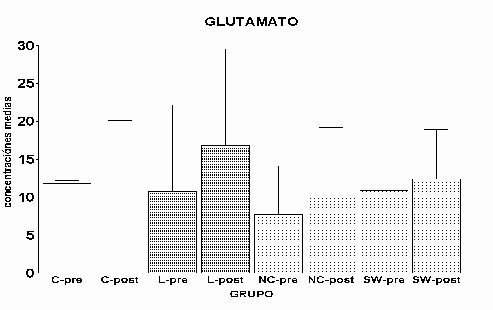
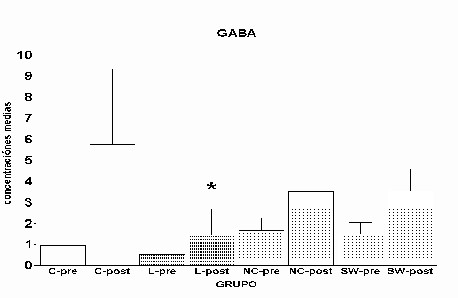
Fig. 12: Implantes de células de Schwann. Región dorsomedial del estriado ipsi y contralateral al implante. Inmunomarcaje para GFAP, revelado con peroxidasa-DAB. Hemisferio contralateral (1) e implantado (2). Nótese (flecha) la mayor reactividad en este último Análisis global: Para los distintos aminoácidos estudiados se comparó la media de todos los valores del periodo prepulso de los grupos experimentales L, NC y SW con la media del grupo control en el mismo periodo, y la media del periodo postpulso de los grupos experimentales con el mismo periodo del grupo control. En el caso de glutamato y taurina no se observaron diferencias significativas en ninguno de los dos periodos, pre (control) y post (despolarización con K+) (Fig. 12 y Fig. 14)). En el caso de GABA, solo en el periodo postpulso del grupo L se observaron diferencias significativas (p= 0.001) (Fig.13).
Fig.
13: Concentración extracelular de Glutamato. Se muestran
las medias ± la desviación estándar
Fig.
14: Concentración extracelular de GABA. Se muestran las
medias ± la desviación estándar
Fig.
15: Concentración extracelular de Taurina. Se muestran
las medias ± la desviación estándar Las concentraciones extracelulares de los aminoácidos del grupo C se tomaron como referencia- para establecer el nivel basal durante el periodo control (prepulso) y durante el pulso de despolarización con K+ (postpulso). Estos valores se compararon con los del grupo L, que, asumimos representaban en nuestro sistema una máxima perdida de funcionalidad. El glutamato es el principal aminoácido excitador de la vía corticoestriatal (Fonnum, F. y cols., 1981). La destrucción de la vía corticoestriatal implicaría una disminución, en el estriado, de los niveles de glutamato en el grupo L respecto al C, así como una reducción en su liberación, sin embargo, no se apreciaron diferencias estadísticamente significativas entre los grupos L y C a lo largo de todo el experimento (fig. 1 y 13). La respuesta al pulso fue idéntica en ambos grupos, produciéndose un aumento significativo de los niveles extracelulares de éste aminoácido en el periodo de despolarización con respecto al periodo control. Esta aparente contradicción podría ser consecuencia de varios factores, entre ellos, la pobre capacidad de "recaptura" que presenta la fibra de diálisis comparada con la gran efectividad en los mecanismos de recaptación de glutamato. Otro posible efecto que estaría contribuyendo a los bajos niveles de glutamato podría explicarse por el siguiente hecho: la despolarización con K+ provocaría un gran aumento de glutamato, en el espacio extracelular, en un periodo de tiempo muy corto (segundos); como el tiempo de muestreo fue de 15 min., ese aumento puntual se vería diluido en el total de la muestra. Además, podría ocurrir que en el grupo lesionado se hubiera producido un fenómeno parcial de "sprouting" por parte de las neuronas no dañadas, compensando los niveles de liberación de glutamato. El neurotransmisor inhibidor de las "medium spiny neurons" es GABA, éstas reciben señales de terminales sinápticos procedentes de la corteza y envían señales inhibidoras a la sustancia negra directamente, o de una manera indirecta a través del globo pálido y de los núcleos subtalámicos (Gerfen, C.R., 1992). En el grupo L no se produjo un aumento de GABA en respuesta a la despolarización con K+ (fig. 2 y 14). Por el contrario, en los animales control hubo un claro aumento en la liberación de GABA. Una posible explicación a este resultado sería el hecho de que la falta de "input" en una vía provoca que esa vía degenere. En el caso de taurina los niveles en los grupos L y C fueron similares (fig. 3 y 15), y la respuesta al pulso muy significativa. Se sabe que éste aminoácido, presente tanto en la glía como en neuronas, es empleado en el cerebro como osmolito (Solís, J. M. y cols., 1988). Así, la gliosis observada en el grupo L (fig. 4) podría explicar la normalización de los niveles de taurina en este grupo. En el grupo NC, los niveles de glutamato no mostraron diferencias estadísticamente significativas respecto al grupo C (fig. 5 y 13). Sin embargo, el grupo NC mostró una clara tendencia a que éstos niveles fueran menores, lo cual ha de considerarse, ya que podría ser reflejo de un efecto negativo por parte del implante. Pudiendo éste ser el responsable de la gliosis. Los astrocitos reactivos producirían sustancias inhibidoras que afectarían a la regeneración de las fibras dañadas (McKeon, R. J. y cols., 1991; Bovolenta, P y cols., 1993; 1997), y por lo tanto a la liberación de glutamato, multiplicándose de esta forma el efecto de la lesión. Incluso en un nivel anterior podrían afectar al metabolismo e inhibir su síntesis, disminuyendo así la liberación. Por otra parte, bajo el efecto de este tipo de implante, se produjo una liberación de GABA en respuesta al pulso de K+, hecho que no se observó en los animales lesionados (6 y 14). Está descrito que factores procedentes de la glía periférica, como son L1 y N-CAM están implicados en crecimiento y regeneración de neuritas. (Martini, R., 1994). El implante de nervio ciático parece ejercer un efecto positivo en la región DM del estriado, aunque no llega a compensar totalmente el efecto de la lesión, puesto que las concentraciones máximas de GABA son menores que las del grupo C (fig. 6). En el caso de taurina, el implante de nervio ciático no pareció ejercer influencia alguna sobre sus niveles extracelulares, posiblemente al encontrarse almacenado en gran cantidad en la glía su concentración no sufrió alteraciones respecto a la del grupo C (fig. 7 y 15). Los niveles de glutamato en el grupo SW no mostraron diferencias estadísticas respecto al grupo C. Sin embargo, a pesar de la falta de significatividad se observó una tendencia al aumento en los niveles postpulso al compararlo con el grupo NC (fig. 9 y 13). Tal vez en este tipo de implante, procedente de células de Schwann recién diferenciadas, de origen perinatal, concurriesen factores tróficos que podrían explicar este resultado. Dichos factores no estarían presentes en el implante de nervio ciático por proceder de un animal adulto. La liberación de GABA inducida por el pulso despolarizante de K+ alcanzó valores próximos a los descritos en los grupos C y NC (fig. 14). Al igual que en los otros grupos, no se observó ningún efecto sobre los niveles extracelulares de taurina (fig. 15). El implante de células de Schwann provoco una reacción gliótica más notoria que la producida por el implante de NC o la lesión, sobre todo en el estriado dorsomedial, llegándose a detectar un efecto contralateral, que afectó a parte del cuerpo calloso (fig. 12). Una posible explicación a este hecho, tal y como se ha comentado mas arriba, es la presencia en la zona dorsomedial de factores tróficos segregados por este tipo de implante (SW) que por difusión alcanzarían el cuerpo calloso y provocarían la reacción gliótica. Estos factores estarían presentes en el implante de células de Schwann de origen perinatal, y no se encontrarían en el implante de nervio ciático, de origen adulto. Otra posibilidad sería un efecto negativo del implante, que provocaría la muerte de las células, con la consiguiente necrosis y salida al espacio extracelular del contenido citoplasmático, induciendo una hipertrofia de la glía (fig. 12) con objeto de recuperar la homeostasis en la zona.
* Bjorjklund, A., Dunnett, S.B. y Nikkhah, G., (1994). Nigral transplants in the rat Parkinson model: functional limitations and strategies to enhance nigroestriatal reconstructions. en Functional Neural Transplantation. Eds. Dunnett, S.B. y Bjorklund, A. pp 47-70. Raven Press. * Bovolenta, P., Fernaud Espinosa, I., Mendez Otero, R. y Nieto Sampedro, M., (1997). Neurite outgrowth inhibitor of gliotic brain tissue. Mode of action and cellular localization, studied with specific monoclonal antibodies. Eur. J. Neurosci. 9 (5); 977-989. * Bovolenta, P., Wandosell, F.y Nieto Sampedro, M., (1993). Characterization of a neurite outgrowth inhibitor expressed after CNS injury. Eur. J. Neurosci. 5 (5); 454-465. * Brockes, J.P., Fields, K.L. y Raff, M.C., (1979). Studies on cultured rat Schwann cells. I. Establishment of purified populations from cultures of peripheral nerve. Brain Res. 165; 105-118. * Collier, T. J. y Springer, J. E., (1991). Co-grafts of embryonic dopamine neurons and adult sciatic nerve into the denervated striatum enhance behavioral and morphological recovery in rats. Exp. Neurol. 114; 343-350. * Dobrossy, M. D. y Dunnett, S. B., (1997). Unilateral striatal lesions impair response execution on a lateralised choice reaction time task. Behav. Brain Res. 87 (2); 159-171. * Dunnett, S.B., Ryan, C.N., Levin, P.D. Reynolds, M. y Bunch, S.T., (1987a). Functional consequences of embryonic neocortex transplanted to rats with prefrontal cortex lesions. Behav. Neurosci.101, (4); 489-503. * Fonnum, F., Storm Mathisen, J. y Divac, I., (1981). Biochemical evidence for glutamate as neurotransmitter in corticostriatal and corticothalamic fibres in rat brain. Neuroscience. 6 (5); 863-873 *Freed, W. J. y Cannon-Spoor, E., (1989). Cortical lesions interfere with behavioral recovery from unilateral substantia nigra lesions induced by brain grafts. Behav. Brain Res. 32; 279-288. * Gerfen, C. R., (1992). The neostriatal mosaic: multiple levels of compartmental organization. TINS, 15, (4); 133-138. * Herranz, A. S., Cristin, J. L. R., Lerma, J. y Martín del Rio. R., (1985). Incremento de sensibilidad en los análisis por CLAE de los OPA-aminoácidos usando como reactivo de derivación el ácido 3-mercaptopropiónico. Resumenes de la Reunión Científica Anual del Grupo de Cromatografía y Técnicas Afines, R.S.E.Q. pp 58-59, Sevilla. * Herranz, A. S., Heim, R. C., Poltorak, M., Giordano, M. y Freed, W. J.; (1993). Sciatic nerve grafts enhance glutamate uptake in the striatum after cortical lesions. 16th Annual meeting of the European Neuroscience Asociation. Abs. 1169. * Hsu, S.M., Raine, L. y Fanger, H., (1981). Use of Avidin-Biotin-Peroxidase Complex (ABC) in Inmunoperoxidase Techniques. J. Histochem. Cytochem. 29; 577-580. * Isono, M., Geller, H. M., Poltorak, M. y Freed, W. J., (1992). Intracerebral transplantation of the A7 inmortalizad astrocytic cell line. Restorative Neurology and Neuroscience 4; 301-309 * Kesslak, J. P., Nieto-Sampedro, M., Globus, J. y Cotman, C. W., (1986). Transplants of purified astrocytes promote behavioral recovery after frontal cortex ablation. Exp-Neurol. 92 (2); 377-390. * Martini, R., (1994). J. Neurocytol. 23; 1-28. * McKeon, R.J., Schreiber, R.C., Rudge, J.S., y Silver, J., (1991). Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J. Neurosci.. 11 (11); 3398-3411. * Patrissi, G. y Stein, D.G., (1975). Temporal factors in recovery of function after brain damage. Exp. Neurol. 47, 3; 470-480. * Paxinos, G. y Watson, Ch., (1982). The rat brain in stereotaxic coordinates. Academic Press; Sydney. * Solís, J. M., Herranz, A. S., Herreras, O., Lerma, J. y Martín del Río, R., (1988). Does taurine act as osmoragulatory substance in the rat brain ?. Neuroscience Letters. 91; 53-58. |
Si
desea contactar directamente a los autores hágalo
por email a: antonio.sanchez@hrc.es |
|